

中文学名:
蜉蝣





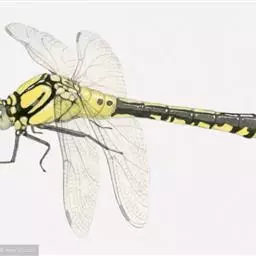
![]() 蜉蝣目(英文:mayfly,学名:Ephemeroptera)通称蜉蝣,具有古老而特殊的形状,是最原始的有翅昆虫。和蜻蜓目可同分为古翅次纲,他们的翅不能折叠。蜉蝣目昆虫体形细长柔软,体长通常为3-27mm,触角短,复眼发达,中胸较大,前翅发达,后翅退化,腹部末端有一对很长的尾须,部分种类还有中央尾丝,稚虫水生,成虫不取食,寿命很短,最短仅一天而已,但它在这短短的生命中,绽放了最绚烂的光彩。
蜉蝣目(英文:mayfly,学名:Ephemeroptera)通称蜉蝣,具有古老而特殊的形状,是最原始的有翅昆虫。和蜻蜓目可同分为古翅次纲,他们的翅不能折叠。蜉蝣目昆虫体形细长柔软,体长通常为3-27mm,触角短,复眼发达,中胸较大,前翅发达,后翅退化,腹部末端有一对很长的尾须,部分种类还有中央尾丝,稚虫水生,成虫不取食,寿命很短,最短仅一天而已,但它在这短短的生命中,绽放了最绚烂的光彩。
基本信息

蜉蝣 蜉蝣目通称蜉蝣,具有古老而特殊的性状,是最原始的有翅昆虫。体形较小或中等,细长,体壁柔软。头部小,触角短,刚毛状。复眼发达,雌性的复眼常左右远离;雄性的复眼常较大,左右接近,且每一复眼上下部小眼面往往不同,一般上半部小眼面大于下半部,也有二部分完全分隔者。单眼3个。口器为咀嚼式,因成虫不取食,没有咀嚼能力,上颚退化消失,下颚也退化,常有下颚须。胸部以中胸最大,亚成虫和成虫都能够在空中飞行。成虫体壁薄而有光泽,常见为白色和淡黄色。有翅一对或两对,飞行时振动频率很小。前、后胸小而不显著,翅有两对,呈三角形,脆弱,膜质,多为前翅大,后翅小,亦有后翅退化者,休息时竖立在身体背面。蜉蝣翅脉相及翅的关节不发达,翅脉最为原始,翅脉极多,多纵脉和横脉,呈网状。翅的表面呈折扇状。足细弱,仅用于攀附。跗节1一5节,末端有爪1对。
腹部11节,第11节仅存窄环形背板。雄性第10节后缘有1对由前足延长形成的抱器,3一4节,少数1节,用于在飞行中抓住雌虫。在其内侧有二对短小简单的阳茎。雌性生殖孔1对,开口于第7、8腹节之腹面。卵巢按节排列。两性生殖孔均成对。腹末有1对分节的长丝状尾须,第11节背板常延长形成中尾丝。尾须和中尾丝细长多节,与缨尾目昆虫相似。
蜉蝣稚虫有两种比较特化的体制:扁平型和鱼型。前者以扁蜉科(Heptageniidae)为代表,虫体扁平,即虫体宽度远大于身体的背腹厚度。胸部的足一般较为宽扁,足的关节转变成前后向,即足一般只能前后运动而不能上下运动,活动时身体腹面与底质不分开,在自然状态下,一般不游泳或游泳能力不强。尾丝上的毛一般散生或环生。鱼型体制以短丝蜉科(Siphlonuridae)、等蜉科(Isonychiidae)以及部分四节蜉科(Baetidae)稚虫为代表。这类蜉蝣的虫体背腹厚度大于虫体的宽度。运动时的体态类似小鱼,即身体呈流线型,足一般细长,中尾丝的两侧和尾须的内侧密生长细毛,相邻的细毛交错成网状,使尾丝具有桨的作用。这类蜉蝣一般可用胸足自由地抓握水中的底质或水生植物,游泳迅速。其它蜉蝣的体制处于这两种之间。
蜉蝣稚虫触角的长度在不同科中变化较大,形态从光滑无毛至密生细毛不等。复眼和单眼变化较小,但细蜉科(Caenidae)中一些种类的单眼表面突出呈棘刺状。另外,唇基、额都可能突出,头顶可能具有各种瘤突和角突。
蜉蝣稚虫的口器是典型的咀嚼式口器,各部分都可能变化,有些变化还很显著。其中以上颚突出呈牙状最为常见。捕食性种类上颚的切齿部分往往发达,而滤食性的种类磨齿部分一般比较发达。
蜉蝣后胸被前翅芽覆盖,背面观不能看见。胸部1~2对翅芽。足的变化较大。蜉蝣总科主要营穴居性生活,足为挖掘足。这种足的腿节和胫节非常粗大,胫节的前侧角突出,爪较小。扁蜉科稚虫足的腿节宽扁,具细毛。
腹部最引人注目的特征是鳃的多样性。鳃的着生位置、对数、大小、形态等各方面都可能变化。腹部背板常具各种不同的刺突和隆起。有些种类腹节背板的侧后角强烈突出并向背方延伸。蜉蝣尾丝的形状多种多样。在活动能力较小的种类中,尾丝各节相对较长而细弱,节上不具毛,只在两节的连接处具稀疏的毛。而游泳能力较强的种类往往中尾丝两侧密生细毛,尾须的内侧长有细毛,相邻的细毛交织成网状而使尾丝具桨的功能,在游泳时产生动力。
蜉蝣稚虫的栖境为方便起见,可将水环境分成两类,一类为静水区,一类为流水区。静水区以湖泊和池塘为代表。进一步可以将静水区光补偿深度以上的区域分成三类,分别为静水水体中、底质表面以及底质中。每类小生境中都有不同的蜉蝣生活。
流水区以溪流和小河为代表,这一栖境也可以分成三类,分别为流水水体中、流水区底质表面及底质缝隙间。

1、静水水体中的自由生活型
这类型的蜉蝣有较大的复眼、较发达的运动肌肉和尾。身体呈流线型,腹部的鳃为膜质片状,单枚或两枚;中尾丝的两侧以及尾丝近体侧密生细毛而使尾丝具桨的功能,与鳃一起击水产生游泳时的动力,也能很好地控制方向。爪一般较细长。当不游泳时,腹部的鳃往往仍然抖动。游泳时足位于腹部并向后沿身体体轴放置,静止时足伸展,能抓握住水中植物。捕食种类以水生的摇蚊和蚊的幼虫为食,也取食水生植物。杂食性的居多。这种类型的典型代表为短丝蜉属(Siphlonurus) 、二翅蜉属(Cloeon)等。
2、静水水体底质表面陷生型
这些蜉蝣一般运动性不强,身体各部分的活动能力不大活动不灵敏,游泳能力不强,采集时要在底质泥沙中寻找,并且要有足够的耐心和细心。身体表面一般具毛,身体一般扁平,腹部的某一对鳃显著扩大而盖住其后的鳃,形成鳃盖。鳃盖在生活时不停地向上抬起,以便于呼吸。鳃盖的边缘往往密生细毛,可以清洁进入鳃盖内的水。尾丝只具有稀疏的环生细毛。生活时陷在靠近水边的泥沙和枯枝落叶的碎屑中,很难发现。个体较小,体色一般为统一的浅色或褐色。少数种类体表具各种瘤突。足向侧方伸展,身体表面常粘附各种泥沙和碎屑。在水中滤食植物或动物碎屑。这一类型的典型代表为细蜉属(Caenis) 、小河蜉属(Potomanthellus)。
3、水区底质中的穴居型
体色一般较淡,体表常具有程度不一的金黄色细毛;头较窄小,额突明显,前缘常分叉;触角的节间常具细毛;上颚牙明显突出于头部前缘,其基部密生细毛;足为挖掘足,位于胸部腹面;腹部的鳃除第1对外,分为两枚,每枚又分为两叉状,缘部呈缨毛状,生活时位于体背,有规律地由前向后运动而整体呈现波动性,能有效地使水在洞穴中流动,以利于滤食。一般生活于沙质或泥质的底质中以逃避敌害。上颚牙用于挖掘时掀起泥土。当这些稚虫从洞中被赶出后,它们立即又进行挖掘新的洞穴。具强烈的避光性。有较好游泳能力,游泳时腹部上下弯曲呈波动形行进。滤食水中碎屑。代表种类为蜉蝣属(Ephemera) 和埃蜉属(Ephoron)。
4、流水区自由生活型
身体苗条,流线型,色彩艳丽。口器各部分密生细毛,触角一般较长。尾桨式。活动迅速,游泳能力强,有趋光性。生活时一般用中后足扒在底质表面,身体略向下倾斜,前足向侧前方伸展,腹部的鳃向外摊开,尾上下击水。前足密生长细毛。这些细毛在前足前缘排列成两行,在细毛之间又夹杂着粗刺。细毛的表面又具有微毛,微毛的顶端呈钩状,相邻的细毛上的微毛相互粘合而使细毛与细毛紧密结合在一起。因此就组成一个筛状的结构。生活时前足前缘上的细毛呈一定角度向前伸展,水流由前向后流动,依靠细毛能有效地过滤水中的碎屑状食物, 代表种类有等蜉属(Isonychia) ,突唇蜉属(Clypeocaenis)。
5、流水区底质表面贴生型
身体一般为浅绿色至褐色。身体扁平,头部非常宽扁。上颚的外侧一般具细毛,下颚须的端部具刷状毛,下唇须表面也具浓密的刷状毛。足的关节呈前后型,因此只能前后向移动。各足的腿节宽扁,胫跗节细长,其表面和后缘往往密生细毛。腹部的鳃侧位。背方的鳃为叶片状,肥厚;腹方的丝状鳃一般位于身体背面,从叶状鳃的基部伸出。各鳃的叶状部分互相叠合成吸盘状,有利于吸附在底质表面。有些种类第1和第7对鳃延伸到身体的腹方,左右两鳃互相接触或接近接触,从而使腹部的鳃整个形成一个吸盘状结构(图3.5)。尾丝上的毛一般较稀疏环生于节间。避光性。刮食石块表面的藻类等食物。能游泳,游泳时主要靠鳃击水。生活状态时可以看到鳃不停地抖动。代表种类为扁蜉科,其中以扁蜉属(Heptagenia)、高翔蜉属(Epeorus)、溪颏蜉属(Rhithrogena) 最典型。
6、流水区底质缝隙栖居型
这种环境中蜉蝣主要有两类。
(1)生活于流水区的水生植物和枯枝落叶中的蜉蝣身体表面往往具各种瘤突或刺,体壁坚硬。腹部背板往往具刺突或脊突。尾丝上常有各种刺。第1对鳃退化,第2或2~3对鳃往往消失。鳃位于身体背面,背方的鳃膜质片状,腹方的鳃分成许多片状小叶。有些种类腹部背板的刺突很大,背板中央强烈隆起,两侧的鳃位于一个腔隙中。生活时一般紧紧抓住水生植物,活动缓慢,采集时常需静候从水生植物中爬出。受到骚扰时,有时将腹部末几节和尾丝向上翻起朝前,然后放直再翻起。游泳时同样如此,整个动作类似蝎子。以撕食性为主。代表种类有小蜉科(Ephemerellidae)的弯握蜉属(Drunella)、锯形蜉属(Serratella) 等。
(2)生活于石块缝隙中的以河花蜉科(Potamanthidae)的种类为代表(Bae & McCafferty 1994)。这类蜉蝣身体一般扁平,为浅黄色至棕红色。上颚具明显的上颚牙,上颚牙上往往具齿和细毛。各足的腿节相对比较宽扁,而胫跗节为细长的圆柱形。前足胫跗节的内缘和背方密生细毛。腹部的鳃侧位,第1对鳃退化,而2~7对鳃的形状与蜉蝣总科的非常类似,但位于身体侧面。腹部背板表面具各种斑纹。尾丝的基部和端部光滑,而中段的两侧具有细毛。游泳动力较强,滤食性。
以上的分类并不严格。每一类的蜉蝣生活环境也非一成不变。其中流水区的种类也往往能在静水区发现,但静水区种类在流水区一般很难发现。总体而言,上述每一类型所选择的栖境仍较严格,采集时在每一种生境中,其代表种类发现概率常是最高的。

幼期(稚虫)水生, 蜉蝣稚虫 生活在淡水湖或溪流中。春夏两季,从午后至傍晚,常有成群的雄虫进行“婚飞”,雌虫独自飞入群中与雄虫配对。产卵于水中。卵微小,椭圆形,具各种颜色,表面有络纹,具粘性,可附着在水底的碎片上。稚虫期数月至1年或1年以上,蜕皮20~24次,多者可达40次。成熟稚虫可见1~2对变黑的翅芽。两侧或背面有成对的气管鳃,是适于水中生活的呼吸器官。吃高等水生植物和藻类,秋、冬两季有些种类以水底碎屑为食。常在静水中攀援、匍匐、或在底泥中潜掘,或在急流中吸附于石砾下栖息。稚虫充分成长后,或浮升到水面,或爬到水边石块或植物茎上,日落后羽化为亚成虫。亚成虫与成虫相似,已具发达的翅,但体色暗淡,翅不透明,后缘有明显的缘毛,雄性的抱握器弯曲不大。出水后停留在水域附近的植物上。一般经24小时左右蜕皮为成虫。这种在个体发育中出现成虫体态后继续蜕皮的现象在有翅昆虫中为蜉蝣目所仅有。这种变态类型特称为原变态。成虫不进食,寿命短,一般只活几小时至数天,所以有“朝生暮死”的说法。蜉蝣成虫在其短暂一生中负责交配,繁衍后代的任务。

蜉蝣亚成虫
变态类型为原变态,一生经历卵、稚虫、亚成虫和成虫4个时期,是有翅亚纲中较原始的变态类型,仅见于蜉蝣目。特别是从幼虫变为成虫要经过一个“亚成虫期”,这时的亚成虫与成虫完全相似。亚成虫期历时较短,一般经数分钟到一天左右即脱皮变为成虫。
雌虫产卵于水中。卵小,白、绿或淡褐色,圆或椭圆形。卵一端常有附着的帽状物,或从卵的表面伸出有粘性的细丝。稚虫水生,常扁平。上颚构造原始,仅一个关节突,中央内缘有一突出的咀嚼面,舌两侧有一对间颚。复眼和单眼发达;触角长,丝状。腹部第1至第7节有成对的气管鳃,尾丝2—3条。其中气管鳃是适于水中生活的呼吸器官,稚虫期一般1一3年,大部分每年1代或两三代,在温暖地区10~15 天能完成发育。一般脱皮20一24次,多者达40次。稚虫取食高等植物和藻类,少数种类捕食水生节肢动物。老熟稚虫一般浮升到水面,或爬到水边石块或植物茎上,日落后羽化成“亚成虫”,此期虫体与成虫相似,但翅不透明或半透明,体表、翅、足常被细毛和缘缨,体色较成虫灰暗,前足和尾须短,行动不活泼,呈静休状态,不能交尾。亚成虫期长短与成虫期之长短有相关现象,亚成虫期短者,成虫寿命则短;亚成虫期长者,成虫期则长。蜉蝣变为成虫后还要脱皮。成虫不取食,寿命极短,只能存活数小时,最多为7天,故有朝生暮死之说。
浮游幼虫包括终生营浮游生活的各类动物的幼体和阶段性浮游生物;后者成体营底栖生活,而幼体是浮游的。浮游幼虫的种类多,数量大,是海洋浮游生物的重要组成部分。到了繁殖旺季,它们在浮游生物中的优势尤为明显,有时整个样品全是幼虫(如蟹类蚤状和大眼幼虫)。除了原生动物以外,几乎所有各类无脊椎动物在发育过程中都经过浮游幼虫阶段,甚至刚孵化出来的仔鱼,因缺乏发达的游泳器官,也只能在海中漂浮,成为浮游幼虫的成员。浮游幼虫的形态也是多种多样的,与成体截然不同。有不少动物的幼虫还历经好几个不同的发育阶段。所以,浮游幼虫是极为复杂的一个生态类群。它们是经济鱼、虾类的天然饵料。因此,它们的数量变化与渔业(尤其是养殖业)的关系十分密切。此外,对附着动物(如藤壶等)的浮游幼虫的附着机制的了解,是制订防除附着生物的方案或增加经济贝类育苗成活率的重要科学依据。

与成虫相比,蜉蝣稚虫的外部形态表现出更加丰富的多样性。这些多样的形态为分类鉴定提供了非常有用的素材。在一些成虫形态变化不大的种类中,如四节蜉科、细蜉科、小蜉科以及河花蜉科,不同种间稚虫的比较对于种类的鉴别常常是非常有效的。
通过对每种生境中代表种类外部形态的了解,根据蜉蝣的外部形态,就可以大体推断出它的生活环境和食性。
由于各种蜉蝣有相对严格的栖境选择,在采集过程中,为了提高采集效率,应该尽可能多地在不同的水生小环境中进行采集取样。同时,要针对各种蜉蝣具有不同的活动能力采取不同的采集方法和技巧。对于那些生活隐蔽且活动能力不强的种类,在采集取样过程中,要有足够的耐心和时间等待它们从隐蔽处出来。而对活动能力非常强的种类,一般很难轻易采到,需要选择拖网或网箱等必要的专门工具和进行相对多次的采集。
由于蜉蝣稚虫形态存在着多样性,可以推知蜉蝣稚虫存在着一定的可塑性,即在自然选择作用下,来自同一祖先的不同支系其稚虫形态可能出现一定的多样性,如细蜉科的稚虫就有三种主要的形态:前足具长毛型、单眼棘刺型和普通型。如果来自不同祖先的若干支系选择同一个生活环境,它们在外部形态上有可能表现出高度的一致性或趋同性。因此,在重建系统发育关系时,稚虫特征的选择应该十分谨慎小心。
由于不同蜉蝣种类及其外部形态与它们的水生生活小环境和生活习性有密切的关系,因此,蜉蝣稚虫在水质监测中得到广泛应用。

1758年Linnaeus 在Systema Naturae第10版中描记了6种蜉蝣,放在蜉蝣属Ephemera中,分为两类,一类具两根尾丝,另一类具有三根尾丝。后来不断有新种报导,Eaton(1883-1888)对前人工作进行了总结,出版了专著A Revisional Monograph of Recent Ephemeridae or Mayflies,共描记了55属300种,将其中的270种分为3 groups(组)9 series(系)14 sections(部)。他的工作奠定了蜉蝣研究的基础,引起了研究的热潮,研究中心逐渐由英国转到了美国,代表人物有Needham(主要是研究北美地区的种类), Navás(全世界),Ulmer(全世界)。1935年,美国的Needham、Traver和中国的徐荫祺先生合著的《蜉蝣生物学》(The Biology of Mayflies)出版,对北美地区的蜉蝣进行了深入而详细的描记和归类,报道蜉蝣共3科17亚科507种。
在这以后,美国的Edmunds、Traver,法国的Demoulin和俄罗斯的Tshernova对该国和一部分世界种类进行报道。1976年,Edmunds et al. 又出版了The mayflies of North and Central America,共列出了中北美地区的蜉蝣共17科24亚科68属622种。在这以后,Edmunds和Peters、McCafferty继续从事这方面的研究。认识比较一致的科约25个。根据Hubbard(1990)统计有310个现存属,61化石属。McCafferty et al.(1989)估计全世界约2 250种。最近几年又陆续有一些报道,全部种类估计不超过2 500种。
关于蜉蝣目在昆虫纲中的系统地位,有许多争论 (Soldán,1997;梁爱萍,1999)。Kukalová-Peck(1968,1978,1983,1985,1994)和Hennig(1981)认为蜉蝣目与蜻蜓目比较接近,与绝灭的古翅类共同组成古翅亚部Palaeoptera,归在有翅部Pterygota中,位置介于衣鱼目Zygentoma与新翅亚部Neoptera之间。
有关目内科级水平的系统发育方面,尽管有一些建议,但均未被普遍接受,有关作者本人的观点也常有改变。已有一些用支序分类方法对某一科内或一些属群的研究;在科级阶元研究方面,声称使用支序方法的就作者所知有四篇(McCafferty & Edmunds,1979;McCafferty,1991;Tomka & Elpers,1991;Kluge,1998),用分析软件演算的工作则尚未见。形成不同观点的主要原因在于各家对一些特征的权值有不同认识,尤其在如何对待和处理成虫和稚虫特征方面认识不同。
在中国,“蜉蝣”作为一个科学动物名词出现在1891年。当时授课于芜湖驿矶山同文书馆的美籍传教士华约翰在《格致汇编》第六年中发表了《虫学略论》,在“分目各论”部分有“第十一蜉蝣类”,绘一虫为例,并加以数行说明(邹树文,1981)。从这篇文章的总体风格和所译名词以及作者的背景来看,它可能不是译自日文。“但其书中所用名词大都不同于现代,加之《格物汇编》主要分销各地领事馆、税务处与礼拜堂,鲜为普通民众所有,因而对中国学术研究影响甚微。”(王思明、周尧,1995)邹树文(1956)说:“日本学者以此名作为Ephemeroptera(蜉蝣目)一目昆虫的名称,中国现今昆虫学上一贯沿用,这是对的。”其实早在1908年沈维钟就写到:“(蜉蝣)实发生于水上,盖其幼虫须在水中一二年,然后变化。变化之后,形态略似蚕蛾,惟尾末有细丝三条,长倍其身。藉之,可以浮游水面。行动如射,故称之。……今日学者所称臼齿蜉蝣是也。……可知蜉蝣断非天牛一类。……”可以看出:沈维钟的描述可能译自日本书;他是第一位作蜉蝣形态描述的中国人,虽然描述十分简单;他明确否定了古人对蜉蝣的一些错误解释。1923年出版的《动物学大辞典》(杜亚泉等编)中的“蜉蝣”条也注有日文。综上所述,邹树文1956年的说法是对的。
中国蜉蝣目的科学研究
第一位以科学的方法报道中国蜉蝣的是英国人Walker。他于1853年发表了Cloeon sinense(当时他命名为Caenis sinense)。接着Eaton (1870—1892)报道了在中国有分布的2种蜉蝣。McLachlan于1904年报道1种。在这以后30年里,中国蜉蝣的研究出现一个高潮。德国人Ulmer(1912—1936)、西班牙传教士Navás(1922—1935)描记了许多新种,随后,徐荫祺先生(Hsu Yin-Qi,1932—1938)对中国的蜉蝣进行了系统研究,胡经甫(1935)在《中国昆虫名录》中统计中国已知蜉蝣41种,去除Navás未定名的,实际是40种。Ulmer(1936)统计中国蜉蝣47种,徐荫祺(1937—1938)统计有64种,并记述5新种。日本人上野益三(Uéno)于1928年报道1种,1931年报道台湾蜉蝣有12种,其中在上述各名录中未见的有Ephemera japonica Mclachlan,Ephemera supposita Eaton、Epeocus psi Eaton、Ecdyonurus hyalinus Ulmer、Rhithrogena vitrea Ulmer、Isonychia formosana Ulmer、Pseudocloeon kraepelini Klapálek共7种。他(1941)还报道了中国东北地区的5种蜉蝣,其中Ephoron virgo(Olivier 1791)和Oligoneuriella rhenana(Imhoff 1852)对当时的中国是新记录种。另外,日本人松村延年(Matsumura,1933)报道1种,另一位日本人今西锦司(Imanishi,1940)也报道了一些中国东北地区的蜉蝣,但大部分未命名。经后人整理和命名,现知有10种过去一直始终未被统计(Bae,1997; Bae & Liu,1999)。去掉异名,在解放前,中国共知蜉蝣88种。在1942—1978年间,国内蜉蝣的研究陷入停顿。在国际上,只有零星的关于中国东北、香港和台湾省、云南省蜉蝣的研究报道,共计6种。Gillies et al.(1949,1951)命名了采自中国香港的Cloeon harveyi和Isca purpurea。1972年,Tshernova 报道中国云南一种Vietnamella ornata,并于1973年报道了Ephemera strigata在中国的分布。1963年,Peters et al. 报道中国香港一种Habrophlebiodes gilliesi,Tsui和Peters于1970年报道了该种的稚虫。上野益三(1969)报道了来自中国台湾和香港地区6种蜉蝣,其中Baetiella japonica(Imanishi)是在中国的首次发现。从60年代开始,徐荫祺先生带领南京师范大学的学生重新进行蜉蝣目的分类研究工作,并于1978年开始发表新种。至今,报道了大量大陆地区的蜉蝣,其中归鸿(1985)对以前的工作进行了统计,共计有96种(去除异名和化石种,实际是92种),并由尤大寿和归鸿(1995)完成了《中国经济昆虫志·蜉蝣目》的编写工作,详细描述了101种蜉蝣。另外韩国人Bae于1991年在研究河花蜉科Potamanthidae时,报道产于中国的2新种。Braasch & Soldán(1980)报道一种。在台湾地区,1981年,Braasch将Ulmer报道分布于台湾的Epeorus psi Eation (1885)改为Epeorus erratus。1985年,Müller-Liebenau报道了台湾四节蜉科Baetidae的6个新种,同年Waltz & McCafferty也报道了一种。康世昌等于1994—1996对台湾地区的蜉蝣分类作了大量工作,发表了46个新种。这些新报道的种除Epeorus erratus外均以稚虫为模式标本。综上所述,根据统计,截止到2001年底,中国已报道蜉蝣256种(表1和表2,部分地方志提及的种类没有统计在内)。Tshernova et al.(1986)提及的中国有分布的种因没有注明具体标本采集地点而没有统计在内。Braasch(1999)报道的中国蜉蝣因在Tshernova et al.(1986)和一些地方志中已有报道,本研究中没有见到标本的均未涉及。

蜉蝣目在昆虫纲中的系统发育地位
关于蜉蝣目在昆虫纲中的系统发育地位有多种观点(详见Kluge 2000)。这些观点基本都是围绕“古翅类”的单系性展开的。“古翅类Palaeoptera”是1922年首次提出的(见Kluge 2000),包括蜉蝣目、蜻蜓目和一些已灭绝的古生代昆虫,与新翅类Neoptera对应。就现存的蜉蝣目与蜻蜓目来看,以下一些特征是古翅类共有和独有的:成虫停息时翅不能折叠覆盖于腹部背面,而只能竖立或平伸于体背;翅具复杂的闰脉;前后翅的形状及脉相是一致的;成虫触角刚毛状;幼期水生;稚虫下颚的内外颚叶愈合,腹部具鳃(全部蜉蝣及部分蜻蜓)等(Hennig,1969;Kukalová-Peck,1991,1997; Wheeler et al.,2001)。然而,以上特征只是古翅类的鉴别特征,不一定是蜉蝣和蜻蜓的共有衍征。对现生所有古翅类共有衍征的不同认识导致三种观点(Hennig,1981;Kristensen,198;Soldán,1997;Wheeler et al,2001)。
第一种观点认为古翅类是一单系群,即蜻蜓和蜉蝣具有一系列共有衍征组成姐妹群。Hennig (1981)提出了它们的四项共有衍征:翅具复杂的闰脉、成虫触角刚毛状、幼期水生、稚虫下颚的内外颚叶愈合。然而,这四项共有衍征中的每一项都有人提出过疑问,其中以Kristensen (1981)的批评最典型:复杂的翅脉很可能为祖征,触角的形态及水生习性平行演化的可能性非常大,而下颚的特征可能不符合蜉蝣的共同模式。
第二种观点认为蜻蜓目与蜉蝣目没有共有衍征,不是姐妹群关系。而是蜉蝣目与(蜻蜓目+新翅类)形成姐妹群关系。Kristensen (1981)举出了蜻蜓目与新翅类七方面的共有衍征:成虫期不蜕皮、通入中后胸的足与翅内的气管来自两个体节、腹部气孔的闭合肌与气孔骨片直接相连、R脉与Rs脉共柄、雌虫的生殖孔单个、中唇舌不发达、头胸部部分肌肉消失。另外,有一系列特征可将蜉蝣目与其它有翅类区别开来(Soldán 1997),如稚虫上颚的滑动关节、腹末具长而分节的中尾丝、翅具肩横脉(subcostal brace 或humeral crossvein brace)以及臀脉A与后肘脉CuP的连接点处形成脉弱点(bulla)、雌虫端滋养式卵巢、精子鞭毛轴丝为9+9+0型(Baccetti et al.,1969)。这种观点也受到部分分子证据的支持(Wheeler 1989; Wheeler et al. 2001)。但是,这一观点受到Kukalová-Peck (1997)逐条批驳,另外这一意见的最主要提倡者Kristensen 的观点在1991年发生了变化。并且,从Wheeler et al. (2001)意见来看,仅从18S rDNA序列得出的结果支持第一种观点,而将形态与分子证据合并分析后,却得出了第二种观点。笔者认为,这可能与形态特征选择有关,因为用于分析的特征是从有关文献中摘录出的,而支持第二种观点的形态特征相对较多。另外,由于现生蜻蜓非常特化(如捕食习性、交尾方式等),自有衍征非常多,有时与其它昆虫很难比较。同时,蜉蝣目具一系列祖征(同时可能也是现生昆虫中仅有的一些特征),它们可能都是一些区别特征,有时很难用于系统发育研究。
第三种观点认为蜉蝣目与新翅类的关系较近,形成姐妹群,它们各自与蜻蜓目的关系较远(Boudreaux 1979,见Kristensen 1981和Wheeler et al. 2001)。Boudreaux(1979)举出了蜉蝣目与新翅类的三个共有衍征:成虫胸部发达的背纵肌、稚虫翅芽的折叠方式、雄虫直接将精子传到雌虫生殖孔。Kristensen对此一一作了评论,指出这些特征要么是祖征(如背纵肌),要么是自有新征(如蜻蜓稚虫的翅芽)或异源同形(如精子的传送方式)。支持这种观点的意见较少。另外,石虫丙、蜉蝣以及蜻蜓的交尾方式都很复杂但形式各异(见郑乐怡、归鸿1999),因此,笔者认为,精子传送方式有待于进一步研究,异源同形的可能极大。
以上三种观点各自都有一些证据,同时也都受到各种批评。仔细分析各观点的支持证据,可以看出,这些特征都是现生昆虫类群所具有的,而现生昆虫大多是高度特化的类群,一些原始特征已经很难辨认。就蜉蝣来看,其成虫翅基骨片已无法辨认。同时,又由于长期进化,许多中间过渡类型绝灭,有翅类内各目之间的特征间隔相对较明显,有时很难进行对比。再者昆虫种类繁多,各种适应型都可能出现,对每种意见都可能找到相反的情况。
在关于古翅类系统发育的研究中,Kukalová-Peck的意见自成体系,需认真考虑、仔细分析。笔者认为,她的观点具有以下一些特点:1)将六足总纲以及节肢动物作为一个整体来考虑,形成统一的体制模式,而不是分割开来各自研究,这就充分考虑到具有共同起源的进化因素;2)将昆虫身体各部分统一到一个共同体节模式中。无论是从古昆虫资料还是现生昆虫特征来看,昆虫身体分部是进化的结果,它们应该有共同的起源;3)充分利用了化石证据。但是,Wheeler et al. (2001)提出Kukalová-Peck的化石证据很难识别。一般而言,对有多年研究经验的专家,化石特征仍然可以识别,只要它们保存得足够好。另外,化石昆虫一般相对较大,如果保存完好,特征可能更易识别。再者,昆虫化石数量很大,可以为研究提供充分的古昆虫特征信息;4)认为现存各种昆虫的器官大多不是新产生的,而是由原始器官演化或退化而来,如蜉蝣稚虫的各种附肢、中尾丝以及头胸部丝状鳃、前翅的肩横脉等的来源及演化在她的体节模式中都能够得到解释;5)考虑到古地理、古气候因素和个体发育信息;6)探讨有翅昆虫各主要类群间的系统发育关系,不可避免地要面对和解决有翅昆虫的起源和演化,而它们的解决有赖于以下五个相关问题的解决:翅的起源和飞行能力的获得、变态起源及各种变态类型的关系、翅脉的演化、翅基骨片及翅位的演化、附肢的演化等。在Kukalová-Peck 的原始昆虫模式中,以上几方面都能得到相对统一的解释。
Kukalová-Peck (1978,1983,1985,1991)提出一个昆虫纲的原始模式,并于1997年对其作了进一步的说明,支持古翅类是单系群的观点。她(1978)指出,用来限定或区别古翅类与新翅类的三个主要特征(翅位、翅面皱褶以及翅基骨片)其实没有可比性,因为现生古翅类与新翅类的翅都是从古生代昆虫原始翅的基础上演化而来,都是衍征,它们是由同一特征朝不同方向演化的结果。而原始模式翅主要特征为:翅至少具8对纵脉,每对脉的前一支为凸脉,后一支为凹脉,凸凹脉相间排列而使翅面呈现皱褶状。在翅基周围具有一马蹄型环绕的骨片组,每块骨片的结构和形状大体相似。骨片共32块,组成4列8行,每一行对应于一对纵脉。随着进化,这些骨片在蜉蝣目、蜻蜓目以及新翅类中发生了不同的愈合情况。蜉蝣目与蜻蜓目的情况类似,因翅基骨片的退化和愈合而在翅基形成两个大的骨板(platform,它们包含的骨片可能有所不同),而在新翅类翅基骨片愈合得较少,形成了在翅折叠过程起到关键作用的腋片,又有肌肉附着,形成关节而使翅能够折叠(Kukalová-Peck 1997)。虽然古翅类昆虫的翅不能折叠,但与新翅类一样,这种模式仍然是一项进化特征,因此不能认为新翅类的翅是由古翅类的翅演化而来。
她(1997)认为,古翅类与新翅类在以下几方面是有根本分岐的:古翅类前后翅在形态与脉相方面是类似的、M脉具主干、Cu脉的主干始终存在且臀横脉(anal brace)与AA脉连接处膜质化。另外,古翅类的翅基骨片通常规则排列,紧靠Sc至Cu脉的两列翅基骨片愈合,其它脉基部的这些骨片相互紧密绞合在一起。以上除第一项外都是古翅类的自有新征。稚虫下颚的内外颚叶愈合,胸部转节与腹板的愈合不完全以及腹部腹板扩大也是古翅类的自有新征。新翅类的自有新征有:前后翅明显不同,前翅主要起保护作用,窄而厚,后翅主要起飞行作用,薄而宽;与古翅类不同,翅的AA脉分为两组;轭脉区膜质折叠在腹面;后翅臀-轭区扩大,具明显的折叠线,臀横脉由骨片及A1组成。蜉蝣与蜻蜓翅脉上的共有衍征有:M脉及Cu脉具主干,CuP脉简单;翅具rp-ma,m-cu,cup-aa1三条重要的横脉;臀横脉长而弯曲。蜉蝣与蜻蜓的共有衍征还有:翅基骨片愈合形成两个骨板而使翅不能折叠、纵脉间具复杂的Y型的闰脉以及臀横脉与CuP连接形成脉弱点(蜉蝣)或翅结(蜻蜓)(Kukalová-Peck 1997)。
后分子系统学为这一研究提供了新信息。Wheeler (1989)根据rDNA以及Wheeler et al. (2001)综合基因序列和形态特征认为蜉蝣目、蜻蜓目与新翅类的关系为蜉蝣目+(蜻蜓目+新翅类)。但是,Hovm Hller et al.(2002)根据更多类群的基因序列分析,提出古翅类是单系群。由于现今所知的有关古翅类基因的信息量还很少(尤其是蜉蝣,见GenBank),很难判断孰优孰劣,因此分子证据有待进一步检验。
笔者认为,解决古翅类的系统发育问题,首先要解决的是Kukalová-Peck的昆虫模式问题,而蜉蝣和蜻蜓是极好的实验材料。如果如她所述,现生蜉蝣的亚成虫期是原始多个龄期的集中或遗迹,不知是否还存在着有两个或多个“亚成虫期”的蜉蝣种类或昆虫。也许对南美、澳大利亚或东南亚地区蜉蝣种类的深入研究会出现另人振奋的结果。另外根据Kukalová-Peck的原始模式,昆虫胸腹部各节都具一对翅(或鳃,或类似结构)和一对附肢。在个别蜉蝣稚虫,其胸部三节各具一对翅芽或其残迹(如Ecdyonurus,Dolania属),另具1对足。但在现生蜉蝣和蜻蜓中,就笔者所知,还没有发现腹部同一体节同时具附肢和鳃的种类。不知以后的研究能否发现这种类型的昆虫。自然界中存在过这种昆虫吗? 如果存在过,现今还存在吗? 在不同的蜉蝣种类中,稚虫的下颚、下唇、三对足的基部可能各具一簇丝状鳃(Sys & Soldan 1980;Zhou & Peters 2003;Muller 1985)。这些鳃支持了Kukalová-Peck的昆虫模式。更多原始昆虫的发现可以为古翅类的研究提供直接有力的证据。
化石证据在Kukalová-Peck原始模式中起到关键作用。更完好的化石发现可能为古翅类的系统发育研究提供新的佐证。分子证据现仍很不够。随着更多古翅类基因序列和线粒体基因组的测定,可以为古翅类的研究找到更多更肯定的分子证据。
虽然蜉蝣目在有翅昆虫中的确切的系统发育地位仍无定论,但一个基本的事实可以肯定,即蜉蝣是无翅昆虫与有翅昆虫之间过渡的纽带,是有翅昆虫中较原始的类群。

“蜉蝣”一词最早出现于《诗经·国风·曹》中的《蜉蝣》诗(可能作于曹昭公在位的公元前661—公元前651年间)。其诗云:
(序)《蜉蝣》,刺奢也。昭公国小而迫,无法自守。好奢而任小人,将无所归依焉。
蜉蝣之羽,衣裳楚楚。心之忧矣,于我归处?
蜉蝣之翼,采采衣服。心之忧矣,于我归息?
蜉蝣掘阅,麻衣如雪。心之忧矣,于我归说?
诗中最难解的是最后一句。根据邹树文(1956)的考证,“掘阅”是双声叠韵,同“阅”,即容貌,并且与上两句中的“之羽”、“之翼”对应。而蓝菊荪(1982)和陈子展(1983)都将此解释为“改变容貌”,多出一个动词,似乎没有邹树文的解释好。他们两人对诗中最后一个字“说”的解释为“歇”。
从诗序和诗文内容来看,诗人是借漂亮而短命的蜉蝣来讽刺时事,表达朝不保暮的忧心。可见诗中的“蜉蝣”即是现今昆虫纲Ephemeroptera目的昆虫,故将本目译为“蜉蝣目”是对的(邹树文,1956,1981)。
由于历代对《诗经》的解释层出不穷,故古代许多人对蜉蝣都有过记述和考证,但可惜绝无具体描述形态的。
《尔雅》可能始创于西周(公元前1027始)。自其创始,直到西汉,历时在千年以上,其间不断有所增补。其在“说虫”部说:“蜉蝣渠略”。
荀况(公元前三世纪人)《荀子·大略篇》:“不饮不食者,蜉蝣也。”
毛亨(公元前二世纪人)《毛传》:“蜉蝣,渠略也。朝生暮死,犹有羽翼以自修饰。楚楚,鲜明貌。采采,众多也。掘阅,容阅也。如雪,言鲜洁。”
西汉戴德(约公元前93—49)《夏小正》:“五月蜉蝣有殷。殷,众也。蜉蝣殷之时也。蜉蝣者,渠略也,朝生而暮死,有何也,有见也。”
三国魏国阮籍(公元三世纪初)《咏怀》:“蜉蝣玩三朝,采采修羽翼。”
晋朝郭义恭(公元三世纪人)《广志》:“蜉蝣可烧啖,美于蝉。蜉蝣在水中翕然生,覆水上,寻死,随流而去。”
晋朝傅咸(公元239—294)《蜉蝣赋》:“读诗至蜉蝣,感其虽朝生暮死,而能修其羽翼,可以有兴,遂赋之。有生之薄,是曰蜉蝣。育微微之陋质,羌采采而自修。不识晦朔,无意春秋。取足一日,尚又何求?戏渟淹而委余,何必江湖而是游。”
唐张九龄《感怀》:“鱼游乐深池,鸟栖欲高枝,嗟尔蜉蝣羽,薨薨亦何为?”
宋苏轼《前赤壁赋》(公元1082年):“寄蜉蝣于天地,渺沧海之一粟。”
宋罗愿(公元1174)《尔雅翼·释虫》:“蜉蝣朝生而暮死。盖蜉蝣者速死之物,故以刺曹公之好奢,言虽衣服楚楚,安能久也。淮南子曰,蚕食而不饮,二十二日而化。蝉饮而不食,三十日而蜕。蜉蝣不食不饮,三日而死。又曰鹤寿千岁以极其游,蜉蝣朝生而暮死尽其乐。盖以旦暮为期,远不过三日尔。蜉蝣出有时,故王褒颂圣主得贤臣颂云,蟋蟀俟秋吟,蜉蝣出以阴,言知时也。又许叔重注《淮南子》言,朝菌者,朝生暮死之虫也。生水上,状似蚕蛾,一名孳母,海南谓之虫邪。则亦蜉蝣之类。按今水上有虫,羽甚整,白露节后即群浮水上,随水而去,以千百计。宛陵人谓之白露虫。” 明刘昌:《县笥琐探恃才傲物》:“汤家公子喜夸诩,好似蜉蝣撼大树。”
另外,清陈梦雷等在《古今图书集成·禽虫典》(公元1726年)和日本人冈元凤(公元1784)在《毛诗品物图考》中还绘出了蜉蝣图。
由上可知,历代古人对蜉蝣外观和生活习性的观察是很细微的,清楚指明蜉蝣是一种生活期极短的昆虫,生活在水中,死后随水漂流。蜉蝣外表漂亮但很柔弱。可见古人的观察是符合实际的。
但同时,一部分古人可能因讲不同方言的缘故,而将“蜉蝣”错误地解释为一类甲虫或蛾子,故朱弘复、高金声(1950)仍认为蜉蝣可能是一种金龟子。如:
三国吴人陆玑(公元三世纪初)《毛诗草木虫鱼疏》:“蜉蝣,方土语也。通谓之渠略。似甲虫,有角。大如指,长三四寸。甲下有翅能飞。夏月阴雨时地中出。今人烧炙之,美如蝉也。”
晋郭璞(公元四世纪初)《尔雅注》:“(蜉蝣)似虫吉虫羌,身狭而长,有角,黄黑色。聚生粪土中,朝生暮死。猪好啖之”。《游仙诗》:“借问蜉蝣辈,宁知龟鹤年。”他在《尔雅音图》中还绘出了蜉蝣图。
孔颖达(公元642年)《毛诗正义》:“郭璞曰(蜉蝣)似虫吉虫羌,身狭而长,有角,黄黑色。聚生粪土中,朝生暮死。猪好啖之。陆玑疏云,蜉蝣,方土语也。通谓之渠略。似甲虫,有角。大如指,长三四寸。甲下有翅能飞。夏月阴雨时地中出。今人烧炙之,美如蝉也。樊光谓之粪中蝎虫,随阴雨时为之朝生暮死。”
北宋陆佃《土卑雅·释虫》(公元1125年其子作序):“(蜉蝣)虫似天牛能飞。烧而啖之,美如蝉也。翕然生。覆水上,寻死,随流。”
南宋戴侗(公元1241—1276)《六书故》:“按诗称蜉蝣羽翼以兴,衣服之采采粲粲,甲虫虽有羽毛,咏物者不应舍甲而言羽。荀子言蜉蝣不饮不食,渠略攻食粪土,亦非不饮不食者也。蜉蝣盖羽虫,疑即夜蛾。夜蛾粉翅,如蝶而小,四月阴雨时多生麦田中,夜则飞扑灯火。不饮食类蚕蛾,故其不能久,近似所谓朝生暮死者。盖以其不饮食而但翻飞,故谓之蜉蝣也邪。”
明李时珍(公元1518—1593)《本草纲目·虫羌虫良 条·附录蜉蝣》(公元1587):“蜉蝣一名渠略。似虫吉虫羌而小,身狭而长,有角,黄黑色,甲下有翅能飞。夏月雨后丛生粪土中,朝生暮死。猪好啖之。人取炙食,云美如蝉也。盖虫羌螂、蜉蝣、腹虫育、天牛,皆蛴螬、蠹、蝎所化此。亦虫羌虫良 之一种,不可不知也。或曰蜉蝣水虫也,状似蚕蛾。朝生暮死。”
明王土斤、王恩义父子(公元1609年)《三才图会》:“(蜉蝣)形似天牛而小,有角,长三四寸,黄黑色,早下有翅能飞。烧而啖之,美如蝉也。翕然生。覆水上,寻死,随流。梁宋之间曰渠略。丛生粪土中,朝生暮死。有浮游之义,故曰蜉蝣也。”
蜉蝣是中国文人骚客心仪的对象。《诗经·曹风》就歌唱过:「蜉蝣之羽,衣裳楚楚……蜉蝣之翼,采采衣服」,把蜉蝣羽翼同妇女衣裙联系起来了,像轻云舒卷,如嫩柳拂水。《淮南子》进一步说:「蚕食而不饮,二十二日而化;蝉饮而不食,三十日而蜕;蜉蝣不食不饮,三日而死」,更作了科学上的比较描述。到明朝李时珍的《本草纲目》,更加了神来的一笔:「蜉,水虫也,……朝生暮死」,一句话抓住了蜉蝣的生态特征。西洋人也早就发现了蜉蝣夭寿,它的昆虫学学名叫做ephemeron,是希腊哲人亚理士多德给起的,意思直截了当就是「短促」。
在五月天阿信的词中,“大时代你我都是蜉蝣”其意,在短暂而有限的生命中,绽放人生的光彩。

2013年8月25日夜间,匈牙利多瑙河沿岸突现数百万只蜉蝣飞虫。弥漫在空气中的飞虫要么黏在行人的脸上,要么爬满车身。第二天清晨,只见地面上布满雄性蜉蝣飞虫的尸体,而其余的则在一夜之间消失不见。这种现象是近40年以来首次出现,令过往行人大为震惊。
2014年7月20日晚间8点45分,密西西比河流域爆发大规模蜉蝣昆虫入侵。团团昆虫聚集在一起酷似“黑云”,并导致三车相撞。它们遮天蔽日,甚至还惊动了该区域的气象雷达。
这些蜉蝣暴风雨般出现在美国威斯康辛州拉克罗斯河域。它们顺着北风迅速向密西西比河上游移动,并出现在雷达屏幕上,就像该区域出现了中轻度降雨。大约三小时之后,随着蜉蝣扩散开来,雷达图像逐渐消失 。
据英国《每日电讯报》6月15日报道,日前,美国宾夕法尼亚州(夏令时UTC-4,比北京时间晚12小时)东部的一座大桥上惊现大量蜉蝣,场面似6月飞雪,已经引发多起交通事故,警方不得不连夜封锁大桥 。
蜉蝣是最原始的有翅昆虫之一。当地时间6月13日晚(北京时间14日上午),一座大桥上出现大量蜉蝣,导致能见度降低。蜉蝣一窝蜂地围着路灯转,最后落在桥面上,堆成厚厚的一层,使得路面光滑不堪,引发多起交通事故。
警方连夜出动警力封锁大桥,6月14日早晨(北京时间14日傍晚)才重新开放。消防队长称,当晚的情景好似六月飞雪,路面也光滑似冰面,共引发3起摩托车事故。

1.又到蜉蝣婚飞时[图]
2015年06月20日00:43 来源:科技日报
6月18日,在匈牙利中部的蒂绍屈尔特,一名男子拍摄蒂萨河里的长尾蜉蝣。
每年的春夏之交,蒂萨河上的大量蜉蝣羽化成虫,婚飞交尾,场面蔚为壮观。短命的成虫只有几个小时的时间进行交尾,然后凄然死亡。雌虫的卵沉入河底后,幼虫在河底会呆长达3年的时间,然后才能变为成虫。
2.解密长尾蜉蝣的短暂一生
长尾蜉蝣短暂一生:成熟化蝶到死亡仅数小时
每年的夏季,也是蒂萨河“盛开”的季节。数以百万计的长尾蜉蝣似乎是瞬间出现于河面之上,整个场景相当壮观。这种昆虫从成熟化蝶到繁殖,最后再到死亡,一生仅仅持续数小时。千上万的长尾蜉蝣会活跃于东欧蒂萨河上,形成了“盛开的蒂萨河”的壮观景象。
在幼虫阶段,长尾蜉蝣要在水下生活长达三年左右的时间。
长尾蜉蝣在孵化后短短数小时即结束生命前争相交配繁殖。
在布达佩斯附近的蒂萨河上,一只长尾蜉蝣正在寻找交配对象。

白蚁是等翅目白蚁科的昆虫类动物。体形较小,繁殖蚁两对翅膀长度超过身体,…





鳞翅目斑蝶科的昆虫,有时亦指蛱蝶科斑蝶亚科的蛱蝶。著名者如帝王蝶及后蝶。多产于热带,…





埃及螳螂是一种螳螂,广泛分布于一些非洲国家…





寄蝇,是双翅目寄蝇科昆虫。已知寄蝇类昆虫共有4亚科37族257属1109种,占世界已知总种数的11%,…





腹长75-78毫米;后翅长62-68毫米。头部黑色,复眼褐色,略带绿色。前额顶小斑,…




蝶角蛉是Ascalaphidae:属于昆虫纲Insecta,脉翅目Neuroptera,…





竹节虫,竹节鞭,属有翅亚纲下的直翅总目,因身体修长而得名,有翅或无翅。体长而大,为中型或大型昆虫,…





九香虫是蝽科九香虫属昆虫九香虫的全虫。全体扁椭圆形,紫黑色,带有铜色光泽;头部狭尖,…

















































